| Dinosaurs Temporal range: Late Triassic–Late Cretaceous, 231.4–65.5 Ma Descendant taxon Aves survives to present | |
|---|---|
 | |
| Mounted skeletons of Tyrannosaurus (left) and Apatosaurus (right) at the American Museum of Natural History | |
| Scientific classification [ e ] | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| clade: | Dinosauriformes |
| Superorder: | Dinosauria Owen, 1842 |
| Orders and suborders | |
Dinosaurs are a diverse and varied group of animals; birds, at over 9,000 species, are the most diverse group of vertebrate besides perciform fish.[3] Paleontologists have identified over 500 distinct genera[4] and more than 1,000 different species of non-avian dinosaurs.[5] Dinosaurs are represented on every continent by both extant species and fossil remains.[6] Some dinosaurs are or were herbivorous, others carnivorous. Some have been, and all extant dinosaurs are, bipedal, others quadrupedal, and others have been able to shift between these body postures. Many non-avian species developed elaborate skeletal modifications such as bony armor, horns or crests. Avian dinosaurs have been the planet's dominant flying vertebrate since the extinction of the pterosaurs. Although generally known for the large size of some species, most dinosaurs were human-sized or even smaller. Most groups of dinosaurs are known to have built nests and laid eggs.
The term "dinosaur" was coined in 1842 by the English paleontologist Richard Owen, and derives from Greek δεινός (deinos) "terrible, powerful, wondrous" + σαῦρος (sauros) "lizard". Through the first half of the 20th century, most of the scientific community believed dinosaurs to have been sluggish, unintelligent cold-blooded animals. Most research conducted since the 1970s, however, has indicated that dinosaurs were active animals with elevated metabolisms and numerous adaptations for social interaction.
Since the first dinosaur fossils were recognized in the early 19th century, mounted dinosaur skeletons have been major attractions at museums around the world, and dinosaurs have become a part of world culture. They have been featured in best-selling books and films such as Jurassic Park, and new discoveries are regularly covered by the media. In informal speech, the word "dinosaur" is used to describe things that are impractically large, obsolete, or bound for extinction,[7] reflecting the outdated view that dinosaurs were maladapted monsters of the ancient world.[citation needed]
Contents[hide] |
Name
The taxon Dinosauria was formally named in 1842 by Sir Richard Owen, who used it to refer to the "distinct tribe or sub-order of Saurian Reptiles" that were then being recognized in England and around the world.[8]:103 The term is derived from the Greek words δεινός (deinos meaning "terrible", "powerful", or "wondrous") and σαῦρος (sauros meaning "lizard" or "reptile").[8]:103[9] Though the taxonomic name has often been interpreted as a reference to dinosaurs' teeth, claws, and other fearsome characteristics, Owen intended it merely to evoke their size and majesty.[10]Modern definition

Under phylogenetic taxonomy, dinosaurs are usually defined as the group consisting of "Triceratops, Neornithes [modern birds], their most recent common ancestor, and all descendants".[11] It has also been suggested that Dinosauria be defined with respect to the MRCA of Megalosaurus and Iguanodon, because these were two of the three genera cited by Richard Owen when he recognized the Dinosauria.[12] Both definitions result in the same set of animals being defined as dinosaurs, that is "Dinosauria = Ornithischia + Saurischia", which encompasses theropods (mostly bipedal carnivores and birds), ankylosaurians (armored herbivorous quadrupeds), stegosaurians (plated herbivorous quadrupeds), ceratopsians (herbivorous quadrupeds with horns and frills), ornithopods (bipedal or quadrupedal herbivores including "duck-bills"), and presumptuously, sauropodomorphs (mostly large herbivorous quadrupeds with long necks and tails).
Many paleontologists note that the order in which sauropodomorphs and theropods diverged may omit sauropodomorphs from the definition for both saurischians and dinosaurs. To avoid the instability of Dinosauria, a more conservative definition of Dinosauria is defined with respect to four anchoring nodes: Triceratops horridus, Saltasaurus loricatus, and Passer domesticus, their most recent common ancestor, and all descendants. This "safer" definition can be expressed as "Dinosauria = Ornithischia + Sauropodomorpha + Theropoda".[13]
There is a wide consensus among paleontologists that birds are the descendants of theropod dinosaurs. Using the strict cladistical definition that all descendants of a single common ancestor must be included in a group for that group to be natural, birds would thus be dinosaurs and dinosaurs are, therefore, not extinct. Birds are classified by most paleontologists as belonging to the subgroup Maniraptora, which are coelurosaurs, which are theropods, which are saurischians, which are dinosaurs.[14]
From the point of view of cladistics, birds are dinosaurs, but in ordinary speech the word "dinosaur" does not include birds. Additionally, referring to dinosaurs that are not birds as "non-avian dinosaurs" is cumbersome. For clarity, this article will use "dinosaur" as a synonym for "non-avian dinosaur". The term "non-avian dinosaur" will be used for emphasis as needed.
General description

Dinosaurs were an extremely varied group of animals; according to a 2006 study, over 500 dinosaur genera have been identified with certainty so far, and the total number of genera preserved in the fossil record has been estimated at around 1850, nearly 75% of which remain to be discovered.[4] An earlier study predicted that about 3400 dinosaur genera existed, including many which would not have been preserved in the fossil record.[19]As of September 17, 2008, 1047 different species of dinosaurs have been named.[5] Some were herbivorous, others carnivorous. Some dinosaurs were bipeds, some were quadrupeds, and others, such as Ammosaurus and Iguanodon, could walk just as easily on two or four legs. Many had bony armor, or cranial modifications like horns and crests. Although known for large size, many dinosaurs were human-sized or smaller. Dinosaur remains have been found on every continent on Earth, including Antarctica.[6] No non-avian dinosaurs are known to have lived in marine habitats or in aerial habitats, although it is possible some feathered non-avian theropods were flyers. There is also evidence that some spinosaurids had semi-aquatic habits.[20]
Distinguishing anatomical features
While recent discoveries have made it more difficult to present a universally agreed-upon list of dinosaurs' distinguishing features, nearly all dinosaurs discovered so far share certain modifications to the ancestral archosaurian skeleton. Although some later groups of dinosaurs featured further modified versions of these traits, they are considered typical across Dinosauria; the earliest dinosaurs had them and passed them on to all their descendants. Such common features across a taxonomic group are called synapomorphies.A detailed assessment of archosaur interrelations by S. Nesbitt[21] confirmed or found the following 12 unambiguous synapomorphies, some previously known:
- in the skull, a supratemporal fossa (excavation) is present in front of the supratemporal fenestra
- epipophyses present in anterior neck vertebrae (except atlas and axis)
- apex of deltopectoral crest (a projection on which the deltopectoral muscles attach) located at or more than 30% down the length of the humerus (upper arm bone)
- radius shorter than 80% of humerus length
- fourth trochanter (projection where the caudofemoralis muscle attaches) on the femur (thigh bone) is a sharp flange
- fourth trochanter asymmetrical, with distal margin forming a steeper angle to the shaft
- on the astragalus and calcaneum the proximal articular facet for fibula occupies less than 30% of the transverse width of the element
- exocciptials (bones at the back of the skull) do not meet along the midline on the floor of the endocranial cavity
- proximal articular surfaces of the ischium with the ilium and the pubis separated by a large concave surface
- cnemial crest on the tibia (shinbone) arcs anterolaterally
- distinct proximodistally oriented ridge present on the posterior face of the distal end of the tibia


Natural history
Origins and early evolution
Main article: Evolution of dinosaurs

When dinosaurs appeared, terrestrial habitats were occupied by various types of basal archosaurs and therapsids, such as aetosaurs, cynodonts, dicynodonts, ornithosuchids, rauisuchias, and rhynchosaurs. Most of these other animals became extinct in the Triassic, in one of two events. First, at about the boundary between the Carnian and Norian faunal stages (about 215 million years ago), dicynodonts and a variety of basal archosauromorphs, including the prolacertiforms and rhynchosaurs, became extinct. This was followed by the Triassic–Jurassic extinction event (about 200 million years ago), that saw the end of most of the other groups of early archosaurs, like aetosaurs, ornithosuchids, phytosaurs, and rauisuchians. These losses left behind a land fauna of crocodylomorphs, dinosaurs, mammals, pterosaurians, and turtles.[11]

Low diversification in the Cretaceous
Statistical analyses based on raw data suggest that dinosaurs diversified, that is, the number of species increased, in the Late Cretaceous. However in July 2008 Graeme T. Lloyd et al. argued that this apparent diversification was an illusion caused by sampling bias, because Late Cretaceous rocks have been very heavily studied. Instead, they wrote, dinosaurs underwent only two significant diversifications in the Late Cretaceous, the initial radiations of the euhadrosaurs and ceratopsians. In the Mid Cretaceous, the flowering angiosperm plants became a major part of terrestrial ecosystems, which had previous been dominated by gymnosperms such as conifers. Dinosaur coprolites (fossilized dung) indicate that, while some ate angiosperms, most herbivorous dinosaurs mainly ate gymnosperms. Meanwhile herbivorous insects and mammals diversified rapidly to take advantage of the new type of plant food, while lizards, snakes, crocodilians and birds also diversified at the same time. Lloyd et al. suggest that dinosaurs' failure to diversify as ecosystems were changing doomed them to extinction.[38]Classification
Main article: Dinosaur classification
Dinosaurs (including birds) are archosaurs, like modern crocodilians. Archosaurs' diapsid skulls have two holes, called temporal fenestrae, located where the jaw muscles attach, and an additional antorbital fenestra in front of the eyes. Most reptiles (including birds) are diapsids; mammals, with only one temporal fenestra, are called synapsids; and turtles, with no temporal fenestra, are anapsids. Anatomically, dinosaurs share many other archosaur characteristics, including teeth that grow from sockets rather than as direct extensions of the jawbones. Within the archosaur group, dinosaurs are differentiated most noticeably by their gait. Dinosaur legs extend directly beneath the body, whereas the legs of lizards and crocodilians sprawl out to either side.Collectively, dinosaurs are usually regarded as a superorder or an unranked clade. They are divided into two orders, Saurischia and Ornithischia, depending upon pelvic structure. Saurischia includes those taxa sharing a more recent common ancestor with birds than with Ornithischia, while Ornithischia includes all taxa sharing a more recent common ancestor with Triceratops than with Saurischia. Saurischians ("lizard-hipped", from the Greek sauros (σαυρος) meaning "lizard" and ischion (ισχιον) meaning "hip joint") retained the hip structure of their ancestors, with a pubis bone directed cranially, or forward.[26] This basic form was modified by rotating the pubis backward to varying degrees in several groups (Herrerasaurus,[39] therizinosauroids,[40] dromaeosaurids,[41] and birds[14]). Saurischia includes the theropods (bipedal and mostly carnivores, except for birds) and sauropodomorphs (long-necked quadrupedal herbivores).
By contrast, ornithischians ("bird-hipped", from the Greek ornitheios (ορνιθειος) meaning "of a bird" and ischion (ισχιον) meaning "hip joint") had a pelvis that superficially resembled a bird's pelvis: the pubis bone was oriented caudally (rear-pointing) Unlike birds, the ornithischian pubis also usually had an additional forward-pointing process. Ornithischia includes a variety of herbivores. (NB: the terms "lizard hip" and "bird hip" are misnomers – birds evolved from dinosaurs with "lizard hips".)
-
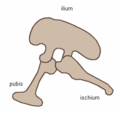 Saurischian pelvis structure (left side)
Saurischian pelvis structure (left side)
-
 Tyrannosaurus pelvis (showing saurischian structure – left side)
Tyrannosaurus pelvis (showing saurischian structure – left side)
-
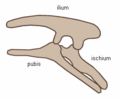 Ornithischian pelvis structure (left side)
Ornithischian pelvis structure (left side)
-
 Edmontosaurus pelvis (showing ornithischian structure – left side)
Edmontosaurus pelvis (showing ornithischian structure – left side)






- Dinosauria
-
- Saurischia (theropods and sauropods)
-
- †Herrerasaurians (early bipedal predators)
- Theropods (all bipedal; most were carnivores)
-
- †Coelophysoids (Coelophysis and close relatives)
- †Ceratosaurians (Ceratosaurus and abelisaurids – the latter were important Late Cretaceous predators in southern continents)
- †Spinosauroids (long bodies; short arms; some with crocodile-like skulls and bony "sails" on their backs)
- †Carnosaurians (Allosaurus and close relatives, like Carcharodontosaurus)
- Coelurosaurians (diverse, with a range of body sizes and niches)
-
- †Tyrannosauroids (small to gigantic, often with reduced forelimbs)
- †Ornithomimosaurians ("ostrich-mimics"; mostly toothless; carnivores to possible herbivores)
- †Therizinosauroids (bipedal herbivores with large hand claws and small heads)
- †Oviraptorosaurians (mostly toothless; their diet and lifestyle are uncertain)
- †Dromaeosaurids (popularly known as "raptors"; bird-like carnivores)
- †Troodontids (similar to dromaeosaurids, but more lightly built)
- Avialans (flying dinosaurs, including modern birds: the only living dinosaurs)
- †Sauropodomorphs (quadrupedal herbivores with small heads, long necks and tails, and elephant-like bodies)
-
- †"Prosauropods" (early relatives of sauropods; small to quite large; some possibly omnivorous; bipeds and quadrupeds)
- †Sauropods (very large, usually over 15 meters long [49 ft])
-
- †Diplodocoids (skulls and tails elongated; teeth typically narrow and pencil-like)
- †Macronarians (boxy skulls; spoon- or pencil-shaped teeth)
-
- †Brachiosaurids (very long necks; forelimbs longer than hindlimbs)
- †Titanosaurians (diverse; stocky, with wide hips; most common in the Late Cretaceous of southern continents)
- †Ornithischians (diverse bipedal and quadrupedal herbivores)
-
- †Heterodontosaurids (meter- or yard-scale herbivores or omnivores with prominent canine teeth)
- †Thyreophorans (armored dinosaurs; mostly quadrupeds)
-
- †Ankylosaurians (scutes as primary armor; some had club-like tails)
- †Stegosaurians (spikes and plates as primary armor)
- †Ornithopods (diverse, from meter- or yard-scale bipeds to 12-meter (39 ft) animals that could move as both bipeds and quadrupeds; evolved a method of chewing using skull flexibility and large numbers of teeth)
-
- †Hadrosaurids ("duckbilled dinosaurs")
- †Pachycephalosaurians ("bone-heads"; bipeds with domed or knobby growth on skulls)
- †Ceratopsians (dinosaurs with horns and frills, although most early forms had only the beginnings of these features)
Evolution and paleobiogeography
Main article: Evolution of dinosaurs
Dinosaur evolution after the Triassic follows changes in vegetation and the location of continents. In the Late Triassic and Early Jurassic, the continents were connected as the single landmass Pangaea, and there was a worldwide dinosaur fauna mostly composed of coelophysoid carnivores and prosauropod herbivores.[42] Gymnosperm plants (particularly conifers), a potential food source, radiated in the Late Triassic. Prosauropods did not have sophisticated mechanisms for processing food in the mouth, and so must have employed other means of breaking down food farther along the digestive tract.[43] The general homogeneity of dinosaurian faunas continued into the Middle and Late Jurassic, where most localities had predators consisting of ceratosaurians, spinosauroids, and carnosaurians, and herbivores consisting of stegosaurian ornithischians and large sauropods. Examples of this include the Morrison Formation of North America and Tendaguru Beds of Tanzania. Dinosaurs in China show some differences, with specialized sinraptorid theropods and unusual, long-necked sauropods like Mamenchisaurus.[42] Ankylosaurians and ornithopods were also becoming more common, but prosauropods had become extinct. Conifers and pteridophytes were the most common plants. Sauropods, like the earlier prosauropods, were not oral processors, but ornithischians were evolving various means of dealing with food in the mouth, including potential cheek-like organs to keep food in the mouth, and jaw motions to grind food.[43] Another notable evolutionary event of the Jurassic was the appearance of true birds, descended from maniraptoran coelurosaurians.[14]By the Early Cretaceous and the ongoing breakup of Pangaea, dinosaurs were becoming strongly differentiated by landmass. The earliest part of this time saw the spread of ankylosaurians, iguanodontians, and brachiosaurids through Europe, North America, and northern Africa. These were later supplemented or replaced in Africa by large spinosaurid and carcharodontosaurid theropods, and rebbachisaurid and titanosaurian sauropods, also found in South America. In Asia, maniraptoran coelurosaurians like dromaeosaurids, troodontids, and oviraptorosaurians became the common theropods, and ankylosaurids and early ceratopsians like Psittacosaurus became important herbivores. Meanwhile, Australia was home to a fauna of basal ankylosaurians, hypsilophodonts, and iguanodontians.[42] The stegosaurians appear to have gone extinct at some point in the late Early Cretaceous or early Late Cretaceous. A major change in the Early Cretaceous, which would be amplified in the Late Cretaceous, was the evolution of flowering plants. At the same time, several groups of dinosaurian herbivores evolved more sophisticated ways to orally process food. Ceratopsians developed a method of slicing with teeth stacked on each other in batteries, and iguanodontians refined a method of grinding with tooth batteries, taken to its extreme in hadrosaurids.[43] Some sauropods also evolved tooth batteries, best exemplified by the rebbachisaurid Nigersaurus.[44]
There were three general dinosaur faunas in the Late Cretaceous. In the northern continents of North America and Asia, the major theropods were tyrannosaurids and various types of smaller maniraptoran theropods, with a predominantly ornithischian herbivore assemblage of hadrosaurids, ceratopsians, ankylosaurids, and pachycephalosaurians. In the southern continents that had made up the now-splitting Gondwana, abelisaurids were the common theropods, and titanosaurian sauropods the common herbivores. Finally, in Europe, dromaeosaurids, rhabdodontid iguanodontians, nodosaurid ankylosaurians, and titanosaurian sauropods were prevalent.[42] Flowering plants were greatly radiating,[43] with the first grasses appearing by the end of the Cretaceous.[45] Grinding hadrosaurids and shearing ceratopsians became extremely diverse across North America and Asia. Theropods were also radiating as herbivores or omnivores, with therizinosaurians and ornithomimosaurians becoming common.[43]
The Cretaceous–Tertiary extinction event, which occurred approximately 65 million years ago at the end of the Cretaceous period, caused the extinction of all dinosaurs except for the birds. Some other diapsid groups, such as crocodilians, lizards, snakes, sphenodontians, and choristoderans, also survived the event.[46]
Paleobiology
Knowledge about dinosaurs is derived from a variety of fossil and non-fossil records, including fossilized bones, feces, trackways, gastroliths, feathers, impressions of skin, internal organs and soft tissues.[47][48] Many fields of study contribute to our understanding of dinosaurs, including physics (especially biomechanics), chemistry, biology, and the earth sciences (of which paleontology is a sub-discipline). Two topics of particular interest and study have been dinosaur size and behavior.Size
Main article: Dinosaur size

Most dinosaurs, however, were much smaller than the giant sauropods. Current evidence suggests that dinosaur average size varied through the Triassic, early Jurassic, late Jurassic and Cretaceous periods.[35] Theropod dinosaurs, when sorted by estimated weight into categories based on order of magnitude, most often fall into the 100 to 1000 kilogram (220 to 2200 lb) category, whereas recent predatory carnivorans peak in the 10 to 100 kilogram (22 to 220 lb) category.[50] The mode of dinosaur body masses is between one and ten metric tonnes.[51] This contrasts sharply with the size of Cenozoic mammals, estimated by the National Museum of Natural History as about 2 to 5 kilograms (5 to 10 lb).[52]
Largest and smallest
Only a tiny percentage of animals ever fossilize, and most of these remain buried in the earth. Few of the specimens that are recovered are complete skeletons, and impressions of skin and other soft tissues are rare. Rebuilding a complete skeleton by comparing the size and morphology of bones to those of similar, better-known species is an inexact art, and reconstructing the muscles and other organs of the living animal is, at best, a process of educated guesswork. As a result, scientists will probably never be certain of the largest and smallest dinosaurs.

Not including modern birds, the smallest known dinosaurs known were about the size of a pigeon.[57] The theropods Anchiornis and Epidexipteryx both had a total skeletal length of under 35 centimeters (1.1 ft).[57][58] Anchiornis is currently the smallest dinosaur described from an adult specimen, with an estimated weight of 110 grams.[58] The smallest herbivorous dinosaurs included Microceratus and Wannanosaurus, at about 60 cm (2 ft) long each.[59][60]
Behavior

The first potential evidence of herding behavior was the 1878 discovery of 31 Iguanodon dinosaurs which were then thought to have perished together in Bernissart, Belgium, after they fell into a deep, flooded sinkhole and drowned.[61] Other mass-death sites have been subsequently discovered. Those, along with multiple trackways, suggest that gregarious behavior was common in many dinosaur species. Trackways of hundreds or even thousands of herbivores indicate that duck-bills (hadrosaurids) may have moved in great herds, like the American Bison or the African Springbok. Sauropod tracks document that these animals traveled in groups composed of several different species, at least in Oxfordshire, England,[62] although there is not evidence for specific herd structures.[63] Dinosaurs may have congregated in herds for defense, for migratory purposes, or to provide protection for their young. There is evidence that many types of dinosaurs, including various theropods, sauropods, ankylosaurians, ornithopods, and ceratopsians, formed aggregations of immature individuals. One example is a site in Inner Mongolia that has yielded the remains of over 20 Sinornithomimus, from one to seven years old. This assemblage is interpreted as a social group that was trapped in mud.[64] The interpretation of dinosaurs as gregarious has also extended to depicting carnivorous theropods as pack hunters working together to bring down large prey.[65][66] However, this lifestyle is uncommon among the modern relatives of dinosaurs (crocodiles and other reptiles, and birds – Harris's Hawk is a well-documented exception), and the taphonomic evidence suggesting pack hunting in such theropods as Deinonychus and Allosaurus can also be interpreted as the results of fatal disputes between feeding animals, as is seen in many modern diapsid predators.[67]


From a behavioral standpoint, one of the most valuable dinosaur fossils was discovered in the Gobi Desert in 1971. It included a Velociraptor attacking a Protoceratops,[75] providing evidence that dinosaurs did indeed attack each other.[76] Additional evidence for attacking live prey is the partially healed tail of an Edmontosaurus, a hadrosaurid dinosaur; the tail is damaged in such a way that shows the animal was bitten by a tyrannosaur but survived.[76] Cannibalism amongst some species of dinosaurs was confirmed by tooth marks found in Madagascar in 2003, involving the theropod Majungasaurus.[77]
Comparisons between the scleral rings of dinosaurs and modern birds and reptiles have been used to infer daily activity patterns of dinosaurs. Although it has been suggested that most dinosaurs were active during the day, these comparisons have shown that small predatory dinosaurs such as dromaeosaurids, Juravenator, and Megapnosaurus were likely nocturnal. Large and medium-sized herbivorous and omnivorous dinosaurs such as ceratopsians, sauropodomorphs, hadrosaurids, ornithomimosaurs may have been cathemeral, active during short intervals throughout the day, although the small ornithischian Agilisaurus was inferred to be diurnal.[78]
Based on current fossil evidence from dinosaurs such as Oryctodromeus, some herbivorous species seem to have led a partially fossorial (burrowing) lifestyle,[79] and some bird-like species may have been arboreal (tree climbing), most notably primitive dromaeosaurids such as Microraptor[80] and the enigmatic scansoriopterygids.[81] However, most dinosaurs seem to have relied on land-based locomotion. A good understanding of how dinosaurs moved on the ground is key to models of dinosaur behavior; the science of biomechanics, in particular, has provided significant insight in this area. For example, studies of the forces exerted by muscles and gravity on dinosaurs' skeletal structure have investigated how fast dinosaurs could run,[82] whether diplodocids could create sonic booms via whip-like tail snapping,[83] and whether sauropods could float.[84]
Communication and vocalization
The nature of dinosaur communication remains enigmatic, and is an active area of research. In 2008, paleontologist Phil Senter examined the evidence for vocalization in Mesozoic animal life, including dinosaurs.[85] Senter found that, contrary to popular depictions of roaring dinosaurs in motion pictures, it is likely that most dinosaurs were not capable of creating any vocalizations. To draw this conclusion, Senter studied the distribution of vocal organs in reptiles and birds. He found that vocal cords in the larynx probably evolved multiple times among reptiles, including crocodilians, which are able to produce guttural roars. Birds, on the other hand, lack a larynx. Instead, bird calls are produced by the syrinx, a vocal organ found only in birds, and which is not related to the larynx, meaning it evolved independently from the vocal organs in reptiles. The syrinx depends on the air sac system in birds to function; specifically, it requires the presence of a clavicular air sac near the wishbone or collar bone. This air sac leaves distinctive marks or opening on the bones, including a distinct opening in the upper arm bone (humerus). While many dinosaurs show evidence of extensive air sac systems, almost none possess the clavicular air sac necessary to vocalize (one exception, Aerosteon, probably evolved its clavicular air sac independently of birds for reasons other than vocalization).[85]The most primitive animals with evidence of a vocalizing syrinx are the enantironithine birds. Any bird-line archosaurs more primitive than this probably did not make vocal calls. Rather, several lines of evidence suggest that dinosaurs used primarily visual communication, in the form of distinctive-looking (and possibly brightly colored) horns, frills, crests, sails and feathers. This is similar to some modern reptile groups such as lizards, in which many forms are largely silent (though like dinosaurs they possess well-developed senses of hearing) but use complex coloration and display behaviors to communicate.[85]
Also, though they may not have been able to vocalize, some dinosaurs may have used other methods of producing sound for communication. Modern animals, including reptiles and birds, use a wide variety of non-vocal sound communication, including hissing, jaw grinding or clapping, use of environment (such as splashing), and wing beating (which would have been possible in winged maniraptoran dinosaurs).[85]
Some studies have suggested that the hollow crests of the lambeosaurines may have functioned as resonance chambers used for a wide range of vocalizations.[86][87] However, Senter (2008) noted that such chambers are also used in modern non-vocal animals to accentuate or deepen non-vocal sounds like hissing. For example, many snakes, which lack vocal cords, have resonating chambers in the skull.[85]
Physiology
Main article: Physiology of dinosaurs

After dinosaurs were discovered, paleontologists first posited that they were ectothermic creatures: "terrible lizards" as their name suggests. This supposed cold-bloodedness implied that dinosaurs were relatively slow, sluggish organisms, comparable to modern reptiles, which need external sources of heat in order to regulate their body temperature. Dinosaur ectothermy remained a prevalent view until Robert T. "Bob" Bakker, an early proponent of dinosaur endothermy, published an influential paper on the topic in 1968.
Modern evidence indicates that dinosaurs thrived in cooler temperate climates, and that at least some dinosaur species must have regulated their body temperature by internal biological means (perhaps aided by the animals' bulk). Evidence of endothermy in dinosaurs includes the discovery of polar dinosaurs in Australia and Antarctica (where they would have experienced a cold, dark six-month winter), the discovery of dinosaurs whose feathers may have provided regulatory insulation, and analysis of blood-vessel structures within dinosaur bone that are typical of endotherms. Skeletal structures suggest that theropods and other dinosaurs had active lifestyles better suited to an endothermic cardiovascular system, while sauropods exhibit fewer endothermic characteristics. It is certainly possible that some dinosaurs were endothermic while others were not. Scientific debate over the specifics continues.[88]

Large dinosaurs would presumably have had to deal with similar issues; their body size suggest they lost heat relatively slowly to the surrounding air, and so could have been what are called inertial homeotherms, animals that are warmer than their environments through sheer size rather than through special adaptations like those of birds or mammals. However, so far this theory fails to account for the numerous dog- and goat-sized dinosaur species, or the young of larger species.
Modern computerized tomography (CT) scans of a dinosaur's chest cavity (conducted in 2000) found the apparent remnants of a four-chambered heart, much like those found in today's mammals and birds.[89] The idea is controversial within the scientific community, coming under fire for bad anatomical science[90] or simply wishful thinking.[91] The question of how this find reflects on metabolic rate and dinosaur internal anatomy may be moot, though, regardless of the object's identity: both modern crocodilians and birds, the closest living relatives of dinosaurs, have four-chambered hearts (albeit modified in crocodilians), and so dinosaurs probably had them as well.[92]
Soft tissue and DNA
One of the best examples of soft-tissue impressions in a fossil dinosaur was discovered in Petraroia, Italy. The discovery was reported in 1998, and described the specimen of a small, very young coelurosaur, Scipionyx samniticus. The fossil includes portions of the intestines, colon, liver, muscles, and windpipe of this immature dinosaur.[47]In the March 2005 issue of Science, the paleontologist Mary Higby Schweitzer and her team announced the discovery of flexible material resembling actual soft tissue inside a 68-million-year-old Tyrannosaurus rex leg bone from the Hell Creek Formation in Montana. After recovery, the tissue was rehydrated by the science team.[48]
When the fossilized bone was treated over several weeks to remove mineral content from the fossilized bone-marrow cavity (a process called demineralization), Schweitzer found evidence of intact structures such as blood vessels, bone matrix, and connective tissue (bone fibers). Scrutiny under the microscope further revealed that the putative dinosaur soft tissue had retained fine structures (microstructures) even at the cellular level. The exact nature and composition of this material, and the implications of Schweitzer's discovery, are not yet clear; study and interpretation of the material is ongoing.[48]
Newer research, published in PloS One (30 July 2008), has challenged the claims that the material found is the soft tissue of Tyrannosaurus. Thomas Kaye of the University of Washington and his co-authors contend that what was really inside the tyrannosaur bone was slimy biofilm created by bacteria that coated the voids once occupied by blood vessels and cells.[93] The researchers found that what previously had been identified as remnants of blood cells, because of the presence of iron, were actually framboids, microscopic mineral spheres bearing iron. They found similar spheres in a variety of other fossils from various periods, including an ammonite. In the ammonite they found the spheres in a place where the iron they contain could not have had any relationship to the presence of blood.[94]
The successful extraction of ancient DNA from dinosaur fossils has been reported on two separate occasions, but, upon further inspection and peer review, neither of these reports could be confirmed.[95] However, a functional visual peptide of a theoretical dinosaur has been inferred using analytical phylogenetic reconstruction methods on gene sequences of related modern species such as reptiles and birds.[96] In addition, several proteins, including hemoglobin,[97] have putatively been detected in dinosaur fossils.[98]
Feathers and the origin of birds
Main article: Origin of birds
The possibility that dinosaurs were the ancestors of birds was first suggested in 1868 by Thomas Henry Huxley.[99] After the work of Gerhard Heilmann in the early 20th century, the theory of birds as dinosaur descendants was abandoned in favor of the idea of their being descendants of generalized thecodonts, with the key piece of evidence being the supposed lack of clavicles in dinosaurs.[100] However, as later discoveries showed, clavicles (or a single fused wishbone, which derived from separate clavicles) were not actually absent;[14] they had been found as early as 1924 in Oviraptor, but misidentified as an interclavicle.[101] In the 1970s, John Ostrom revived the dinosaur–bird theory,[102] which gained momentum in the coming decades with the advent of cladistic analysis,[103] and a great increase in the discovery of small theropods and early birds.[22] Of particular note have been the fossils of the Yixian Formation, where a variety of theropods and early birds have been found, often with feathers of some type.[14] Birds share over a hundred distinct anatomical features with theropod dinosaurs, which are now generally accepted to have been their closest ancient relatives.[104] They are most closely allied with maniraptoran coelurosaurs.[14] A minority of scientists, most notably Alan Feduccia and Larry Martin, have proposed other evolutionary paths, including revised versions of Heilmann's basal archosaur proposal,[105] or that maniraptoran theropods are the ancestors of birds but themselves are not dinosaurs, only convergent with dinosaurs.[106]Feathers
Main article: Feathered dinosaurs

Since the 1990s, a number of additional feathered dinosaurs have been found, providing even stronger evidence of the close relationship between dinosaurs and modern birds. Most of these specimens were unearthed in the lagerstätte of the Yixian Formation, Liaoning, northeastern China, which was part of an island continent during the Cretaceous. Though feathers have been found in only a few locations, it is possible that non-avian dinosaurs elsewhere in the world were also feathered. The lack of widespread fossil evidence for feathered non-avian dinosaurs may be because delicate features like skin and feathers are not often preserved by fossilization and thus are absent from the fossil record. To this point, protofeathers (thin, filament-like structures) are known from dinosaurs at the base of Coelurosauria, such as compsognathids like Sinosauropteryx and tyrannosauroids (Dilong),[108] but barbed feathers are known only among the coelurosaur subgroup Maniraptora, which includes oviraptorosaurs, troodontids, dromaeosaurids, and birds.[14][109] The description of feathered dinosaurs has not been without controversy; perhaps the most vocal critics have been Alan Feduccia and Theagarten Lingham-Soliar, who have proposed that protofeathers are the result of the decomposition of collagenous fiber that underlaid the dinosaurs' integument,[110][111][112]and that maniraptoran dinosaurs with barbed feathers were not actually dinosaurs, but convergent with dinosaurs.[106][111] However, their views have for the most part not been accepted by other researchers, to the point that the question of the scientific nature of Feduccia's proposals has been raised.[113]
Skeleton
Because feathers are often associated with birds, feathered dinosaurs are often touted as the missing link between birds and dinosaurs. However, the multiple skeletal features also shared by the two groups represent another important line of evidence for paleontologists. Areas of the skeleton with important similarities include the neck, pubis, wrist (semi-lunate carpal), arm and pectoral girdle, furcula (wishbone), and breast bone. Comparison of bird and dinosaur skeletons through cladistic analysis strengthens the case for the link.Soft anatomy

Reproductive biology
A discovery of features in a Tyrannosaurus rex skeleton provided evidence of medullary bone in dinosaurs and, for the first time, allowed paleontologists to establish the sex of a dinosaur. When laying eggs, female birds grow a special type of bone between the hard outer bone and the marrow of their limbs. This medullary bone, which is rich in calcium, is used to make eggshells. The presence of endosteally derived bone tissues lining the interior marrow cavities of portions of the Tyrannosaurus rex specimen's hind limb suggested that T. rex used similar reproductive strategies, and revealed the specimen to be female.[117] Further research has found medullary bone in the theropod Allosaurus and the ornithopod Tenontosaurus. Because the line of dinosaurs that includes Allosaurus and Tyrannosaurus diverged from the line that led to Tenontosaurus very early in the evolution of dinosaurs, this suggests that dinosaurs in general produced medullary tissue. Medullary bone has been found in specimens of sub-adult size, which suggests that dinosaurs reached sexual maturity rather quickly for such large animals.[118]Behavioral evidence
Fossils of the troodonts Mei and Sinornithoides demonstrate that some dinosaurs slept with their heads tucked under their arms.[119] This behavior, which may have helped to keep the head warm, is also characteristic of modern birds. Several deinonychosaur and oviraptorosaur specimens have also bee found preserved on top of their nests, likely brooding in a bird-like manner.[120] The ratio between egg volume and body mass of adults among these dinosaurs suggest that the eggs were primarily brooded by the male, and that the young were highly precocial, similar to many modern ground-dwelling birds.[121]Some dinosaurs are known to have used gizzard stones like modern birds. These stones are swallowed by animals to aid digestion and break down food and hard fibers once they enter the stomach. When found in association with fossils, gizzard stones are called gastroliths.[122]
Extinction
Main articles: Cretaceous–Tertiary extinction event and K–T boundary
Non-avian dinosaurs suddenly became extinct approximately 65 million years ago. Many other groups of animals also became extinct at this time, including ammonites (nautilus-like mollusks), mosasaurs, plesiosaurs, pterosaurs, most birds, and many groups of mammals.[6] This mass extinction is known as the Cretaceous–Tertiary extinction event. The nature of the event that caused this mass extinction has been extensively studied since the 1970s; at present, several related theories are supported by paleontologists. Though the consensus is that an impact event was the primary cause of dinosaur extinction, some scientists cite other possible causes, or support the idea that a confluence of several factors was responsible for the sudden disappearance of dinosaurs from the fossil record.At the peak of the Mesozoic, there were no polar ice caps, and sea levels are estimated to have been from 100 to 250 meters (300 to 800 ft) higher than they are today. The planet's temperature was also much more uniform, with only 25 °C (45 °F) separating average polar temperatures from those at the equator. On average, atmospheric temperatures were also much higher; the poles, for example, were 50 °C (90 °F) warmer than today.[123][124]
The atmosphere's composition during the Mesozoic was vastly different as well. Carbon dioxide levels were up to 12 times higher than today's levels, and oxygen formed 32 to 35% of the atmosphere,[citation needed] as compared to 21% today. However, by the late Cretaceous, the environment was changing dramatically. Volcanic activity was decreasing, which led to a cooling trend as levels of atmospheric carbon dioxide dropped. Oxygen levels in the atmosphere also started to fluctuate and would ultimately fall considerably. Some scientists hypothesize that climate change, combined with lower oxygen levels, might have led directly to the demise of many species. If the dinosaurs had respiratory systems similar to those commonly found in modern birds, it may have been particularly difficult for them to cope with reduced respiratory efficiency, given the enormous oxygen demands of their very large bodies.[6]
Impact event
Main article: Chicxulub crater

Although the speed of extinction cannot be deduced from the fossil record alone, various models suggest that the extinction was extremely rapid. The consensus among scientists who support this theory is that the impact caused extinctions both directly (by heat from the meteorite impact) and also indirectly (via a worldwide cooling brought about when matter ejected from the impact crater reflected thermal radiation from the sun).
In September 2007, U.S. researchers led by William Bottke of the Southwest Research Institute in Boulder, Colorado, and Czech scientists used computer simulations to identify the probable source of the Chicxulub impact. They calculated a 90% probability that a giant asteroid named Baptistina, approximately 160 km (100 mi) in diameter, orbiting in the asteroid belt which lies between Mars and Jupiter, was struck by a smaller unnamed asteroid about 55 km (35 mi) in diameter about 160 million years ago. The impact shattered Baptistina, creating a cluster which still exists today as the Baptistina family. Calculations indicate that some of the fragments were sent hurtling into earth-crossing orbits, one of which was the 10 km (6 mi) wide meteorite which struck Mexico's Yucatan peninsula 65 million years ago, creating the Chicxulub crater.[128]
A similar but more controversial explanation proposes that "passages of the [hypothetical] solar companion star Nemesis through the Oort comet cloud would trigger comet showers."[129] One or more of these comets then collided with the Earth at approximately the same time, causing the worldwide extinction. As with the impact of a single asteroid, the end result of this comet bombardment would have been a sudden drop in global temperatures, followed by a protracted cool period.[129]
Deccan Traps
Main article: Deccan Traps
Before 2000, arguments that the Deccan Traps flood basalts caused the extinction were usually linked to the view that the extinction was gradual, as the flood basalt events were thought to have started around 68 million years ago and lasted for over 2 million years. However, there is evidence that two thirds of the Deccan Traps were created in only 1 million years about 65.5 million years ago, and so these eruptions would have caused a fairly rapid extinction, possibly over a period of thousands of years, but still longer than would be expected from a single impact event.[130][131]The Deccan Traps could have caused extinction through several mechanisms, including the release into the air of dust and sulphuric aerosols, which might have blocked sunlight and thereby reduced photosynthesis in plants. In addition, Deccan Trap volcanism might have resulted in carbon dioxide emissions, which would have increased the greenhouse effect when the dust and aerosols cleared from the atmosphere.[131] Before the mass extinction of the dinosaurs, the release of volcanic gases during the formation of the Deccan Traps "contributed to an apparently massive global warming. Some data point to an average rise in temperature of 8 °C (14 °F) in the last half million years before the impact [at Chicxulub]."[130][131]
In the years when the Deccan Traps theory was linked to a slower extinction, Luis Alvarez (who died in 1988) replied that paleontologists were being misled by sparse data. While his assertion was not initially well-received, later intensive field studies of fossil beds lent weight to his claim. Eventually, most paleontologists began to accept the idea that the mass extinctions at the end of the Cretaceous were largely or at least partly due to a massive Earth impact. However, even Walter Alvarez has acknowledged that there were other major changes on Earth even before the impact, such as a drop in sea level and massive volcanic eruptions that produced the Indian Deccan Traps, and these may have contributed to the extinctions.[132]
Failure to adapt to changing conditions
Lloyd et al. (2008) noted that, in the Mid Cretaceous, the flowering, angiosperm plants became a major part of terrestrial ecosystems, which had previously been dominated by gymnosperms such as conifers. Dinosaur coprolite–fossilized dung–indicate that, while some ate angiosperms, most herbivorous dinosaurs ate mainly gymnosperms. Statistical analysis by Lloyd et al. concluded that, contrary to earlier studies, dinosaurs did not diversify very much in the Late Cretaceous. Lloyd et al. suggested that dinosaurs' failure to diversify as ecosystems were changing doomed them to extinction.[38]Possible Paleocene survivors
Main article: Paleocene dinosaurs
Non-avian dinosaur remains are occasionally found above the K–T boundary. In 2001, paleontologists Zielinski and Budahn reported the discovery of a single hadrosaur leg-bone fossil in the San Juan Basin, New Mexico, and described it as evidence of Paleocene dinosaurs. The formation in which the bone was discovered has been dated to the early Paleocene epoch, approximately 64.5 million years ago. If the bone was not re-deposited into that stratum by weathering action, it would provide evidence that some dinosaur populations may have survived at least a half million years into the Cenozoic Era.[133] Other evidence includes the finding of dinosaur remains in the Hell Creek Formation up to 1.3 meters (51 in) above (40000 years later than) the K–T boundary. Similar reports have come from other parts of the world, including China.[134] Many scientists, however, dismissed the supposed Paleocene dinosaurs as re-worked, that is, washed out of their original locations and then re-buried in much later sediments.[135][136] However, direct dating of the bones themselves has supported the later date, with U–Pb dating methods resulting in a precise age of 64.8 ± 0.9 million years ago.[137] If correct, the presence of a handful of dinosaurs in the early Paleocene would not change the underlying facts of the extinction.[135]History of discovery
Dinosaur fossils have been known for millennia, although their true nature was not recognized. The Chinese, whose modern word for dinosaur is konglong (恐龍, or "terrible dragon"), considered them to be dragon bones and documented them as such. For example, Hua Yang Guo Zhi, a book written by Zhang Qu during the Western Jin Dynasty, reported the discovery of dragon bones at Wucheng in Sichuan Province.[138] Villagers in central China have long unearthed fossilized "dragon bones" for use in traditional medicines, a practice that continues today.[139] In Europe, dinosaur fossils were generally believed to be the remains of giants and other creatures killed by the Great Flood.Scholarly descriptions of what would now be recognized as dinosaur bones first appeared in the late 17th century in England. Part of a bone, now known to have been the femur of a Megalosaurus,[140] was recovered from a limestone quarry at Cornwell near Chipping Norton, Oxfordshire, England, in 1676. The fragment was sent to Robert Plot, Professor of Chemistry at the University of Oxford and first curator of the Ashmolean Museum, who published a description in his Natural History of Oxfordshire in 1677. He correctly identified the bone as the lower extremity of the femur of a large animal, and recognized that it was too large to belong to any known species. He therefore concluded it to be the thigh bone of a giant human similar to those mentioned in the Bible. In 1699, Edward Lhuyd, a friend of Sir Isaac Newton, was responsible for the first published scientific treatment of what would now be recognized as a dinosaur when he described and named a sauropod tooth, "Rutellum implicatum",[141][142] that had been found in Caswell, near Witney, Oxfordshire.[143]
Between 1815 and 1824, the Rev William Buckland, a professor of geology at Oxford University, collected more fossilized bones of Megalosaurus and became the first person to describe a dinosaur in a scientific journal.[140][144] The second dinosaur genus to be identified, Iguanodon, was discovered in 1822 by Mary Ann Mantell – the wife of English geologist Gideon Mantell. Gideon Mantell recognized similarities between his fossils and the bones of modern iguanas. He published his findings in 1825.[145][146]

The study of these "great fossil lizards" soon became of great interest to European and American scientists, and in 1842 the English paleontologist Richard Owen coined the term "dinosaur". He recognized that the remains that had been found so far, Iguanodon, Megalosaurus and Hylaeosaurus, shared a number of distinctive features, and so decided to present them as a distinct taxonomic group. With the backing of Prince Albert of Saxe-Coburg-Gotha, the husband of Queen Victoria, Owen established the Natural History Museum in South Kensington, London, to display the national collection of dinosaur fossils and other biological and geological exhibits.
In 1858, the first known American dinosaur was discovered, in marl pits in the small town of Haddonfield, New Jersey (although fossils had been found before, their nature had not been correctly discerned). The creature was named Hadrosaurus foulkii. It was an extremely important find: Hadrosaurus was one of the first nearly complete dinosaur skeletons found (the first was in 1834, in Maidstone, Kent, England), and it was clearly a bipedal creature. This was a revolutionary discovery as, until that point, most scientists had believed dinosaurs walked on four feet, like other lizards. Foulke's discoveries sparked a wave of dinosaur mania in the United States.


After 1897, the search for dinosaur fossils extended to every continent, including Antarctica. The first Antarctic dinosaur to be discovered, the ankylosaurid Antarctopelta oliveroi, was found on Ross Island in 1986, although it was 1994 before an Antarctic species, the theropod Cryolophosaurus ellioti, was formally named and described in a scientific journal.
Current dinosaur "hot spots" include southern South America (especially Argentina) and China. China in particular has produced many exceptional feathered dinosaur specimens due to the unique geology of its dinosaur beds, as well as an ancient arid climate particularly conducive to fossilization.
The "dinosaur renaissance"
Main article: Dinosaur renaissance
The field of dinosaur research has enjoyed a surge in activity that began in the 1970s and is ongoing. This was triggered, in part, by John Ostrom's discovery of Deinonychus, an active predator that may have been warm-blooded, in marked contrast to the then-prevailing image of dinosaurs as sluggish and cold-blooded. Vertebrate paleontology has become a global science. Major new dinosaur discoveries have been made by paleontologists working in previously unexploited regions, including India, South America, Madagascar, Antarctica, and most significantly China (the amazingly well-preserved feathered dinosaurs in China have further consolidated the link between dinosaurs and their conjectured living descendants, modern birds). The widespread application of cladistics, which rigorously analyzes the relationships between biological organisms, has also proved tremendously useful in classifying dinosaurs. Cladistic analysis, among other modern techniques, helps to compensate for an often incomplete and fragmentary fossil record.Cultural depictions
Main article: Cultural depictions of dinosaurs
By human standards, dinosaurs were creatures of fantastic appearance and often enormous size. As such, they have captured the popular imagination and become an enduring part of human culture. Entry of the word "dinosaur" into the common vernacular reflects the animals' cultural importance: in English, "dinosaur" is commonly used to describe anything that is impractically large, obsolete, or bound for extinction.[7]Public enthusiasm for dinosaurs first developed in Victorian England, where in 1854, three decades after the first scientific descriptions of dinosaur remains, the famous dinosaur sculptures were unveiled in London's Crystal Palace Park. The Crystal Palace dinosaurs proved so popular that a strong market in smaller replicas soon developed. In subsequent decades, dinosaur exhibits opened at parks and museums around the world, ensuring that successive generations would be introduced to the animals in an immersive and exciting way.[148] Dinosaurs' enduring popularity, in its turn, has resulted in significant public funding for dinosaur science, and has frequently spurred new discoveries. In the United States, for example, the competition between museums for public attention led directly to the Bone Wars of the 1880s and 1890s, during which a pair of feuding paleontologists made enormous scientific contributions.[149]
The popular preoccupation with dinosaurs has ensured their appearance in literature, film and other media. Beginning in 1852 with a passing mention in Charles Dickens' Bleak House,[150] dinosaurs have been featured in large numbers of fictional works. Sir Arthur Conan Doyle's 1912 book The Lost World, the iconic 1933 film King Kong, 1954's Godzilla and its many sequels, the best-selling 1990 novel Jurassic Park by Michael Crichton and its 1993 film adaptation are just a few notable examples of dinosaur appearances in fiction. Authors of general-interest non-fiction works about dinosaurs, including some prominent paleontologists, have often sought to use the animals as a way to educate readers about science in general. Dinosaurs are ubiquitous in advertising; numerous companies have referenced dinosaurs in printed or televised advertisements, either in order to sell their own products or in order to characterize their rivals as slow-moving, dim-witted or obsolete.[151]
















Tiada ulasan:
Catat Ulasan